



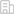

近日,中国农业大学生物学院陈三凤教授课题组在Cell Press细胞出版社旗下的知名学术期刊《生物技术趋势》(Trends in Biotechnology)在线发表了题为《利用合成生物学方法在水稻中表达固氮酶合成途径和克服固氮酶在植物细胞质中的不稳定性》(Using synthetic biology to express nitrogenase biosynthesis pathway in rice and to overcome barriers of nitrogenase instability in plant cytosol)研究论文。
该研究通过多基因组装、单转化、共转化及杂交等技术,将来自2个固氮细菌的13个固氮基因(约40 kb)导入水稻中,水稻基因组重测序表明13个固氮基因整合在水稻1号染色体的2个位点,qRT-PCR和Western blot分析表明固氮基因在水稻中能转录和表达,而且能形成固氮酶的NifDK四聚体,并发现和解决了NifH蛋白在植物细胞质中被蛋白内切酶切割的问题,为实现禾本科植物实现“自主固氮”提供了重要参考。

固氮细菌利用体内的固氮酶在常温和常压下将空气中的N2 还原成氨的过程称为生物固氮 (Biological nitrogen fixation)。禾本科植物由于缺乏根瘤共生固氮体系, 其高产和稳产高度依赖化学氮肥。利用合成生物学方法将固氮酶合成途径直接导入禾本科植物使其实现“自主固氮”, 是有望减少氮肥用量的一个有效途径。
固氮酶是一种非常复杂的金属酶,由2种蛋白组份组成:铁蛋白(也称为NifH蛋白,由 nifH 基因编码)和钼铁蛋白(也称为NifDK蛋白,由 nifD 和 nifK 基因编码)。此外,固氮酶的合成还需要3种金属簇:[Fe4S4] cluster、P-cluster及 FeMo-cofactor [Mo–7 Fe–9S–C–homocitrate]。FeMo-cofactor位于NifDK蛋白上,是N2络合和还原的场所,其他两种金属簇的功能是传递电子。固氮酶的合成需要至少11种固氮基因[ nifH、nifD、nifK、nifB、nifE、nifN、nifV、nifX、nifQ (或 hesA ) 、nifU 及 nifS ]。在植物中异源表达固氮酶面临许多挑战:将多个固氮基因同时导入植物、多个固氮基因在植物中协调表达、以及固氮酶相关蛋白在植物中的稳定性等。
本研究首次将13个固氮酶合成相关基因:来自多粘类芽孢杆菌( Paenibacillus polymyxa )的11个基因( nifH 、 nifD 、 nifK 、 nifX 、 nifE 、 nifB 、 nifN 、 nifV 、 hesA 、 groES 和 groEL )和来自产酸克雷伯菌 ( Klebsiella oxytoca ) 的2个基因( nif U 和 nif S) ,构建在3个植物表达载体,并通过农杆菌介导的单转化和共转化导入水稻中。对筛选获得的T1代转基因水稻进行杂交, 筛选获得含有全部13个固氮基因的第一代(F1)代杂交水稻,再通过水稻连续自交获得了第4代 (F4) 转基因水稻植株。通过PCR和RT-PCR分析, 从256个F4代转基因水稻株系中, 鉴定获得了55个携带全部13个固氮酶合成相关基因的转基因基因水稻株系(图1)。

图1.利用合成生物学方法将13个固氮基因导入水稻的试验方案。阳性植株表示含固氮基因。
随机选取3个F4 转基因水稻株系(L8-17、L12-13及L7-15)进行深入研究。PCR分析表明,13个固氮基因在L8-17株系和L12-13株系中均能稳定遗传,而13个固氮基因在L7-15株系中呈现3:1分离。水稻基因组重测序结果表明,在L8-17株系和L12-13株系中,13个固氮基因均插入到水稻1号染色体的2个位点(图2)。而在L7-15株系中,13个固氮基因插入到水稻2个不同的染色体上。
图2.F4 转基因水稻株系L8-17中13个固氮基因( nif )在水稻1号染色体的2个插入位点及其排列。
(A) 9 个固氮基因 ( nifH nifD nifK nifX hesA nifB nifE nifN nifV ) 在水稻1号染色体上的插入位点及其排列。
(B) 4 个固氮基因 ( nifU nifS groES groEL ) 在水稻1号染色体上的插入位点及其排列。
qRT-PCR分析表明,13个固氮基因在3个F4 转基因水稻株系(L8-17、L12-13及L7-15)中,以不同水平转录(图3A)。Western blot分析表明,11 个Nif蛋白 (NifB、NifH、NifD、NifK、NifE、NifN、NifV、NifX、HesA、NifS 及 GroEL) 在在水稻株系L8-17的细胞质中稳定表达(图3B)。而且,NifD和NifK在水稻细胞质中能形成稳定的NifDK四聚体(图3C),该NifDK四聚体是形成有活性的固氮酶所必需的。同时,发现水稻细胞质中合成的NifH蛋白分子量比固氮细菌P. polymyxa NifH分子量小约1.8 kDa, 推测NifH被植物细胞质中的蛋白酶切割。

图3.F4 转基因水稻中固氮基因(nif )转录和固氮酶蛋白(Nif)表达。
(A) qRT-PCR分析13个固氮基因(nifB nifH nifD nifK nifE nifN nifX hesA nifV nifS nifU groES groEL ) 在3个F4 转基因水稻株系L8-17、L12-13及 L7-15中的转录水平。
(B) Western blot 分析F4 转基因水稻株系L8-17中,11 个Nif 蛋白(NifB、NifH、NifD、NifK、NifE、NifN、NifV、HesA、NifX、 NifS及GroEL)的表达。
(C) Western blot(厌氧和非变性电泳)分析F4 转基因水稻株系 L8-17中NifDK四聚体的合成。
本研究发现在水稻细胞质中表达的NifH 蛋白的分子量变小。为解决NifH在植物细胞质中不稳定性问题,对NifH蛋白氨基酸序列进行以系列缺失和氨基酸替代、以及利用烟草烟瞬时表达系统进行研究,结果表明NifH的切割发生在第17位苏氨酸(T)和第18位丝氨基酸(S)之间。13个NifH氨基酸替代突变体(T17A、T17C、T17I、T17M、T17S、T17V、S18A、S18C、S18G、S18L、S18Q、S18T 及 Q19 A),能使 P. polymyxa Δ nifH 突变株恢复固氮酶活性,特别是3个NifH突变体 (T17C、T17V及S18A) 能使Δ nifH 突变株分别恢复到59.4~71.9%固氮酶活性;烟草瞬时表达也证明这13个NifH氨基酸替代突变体在烟草细胞质中不再被蛋白酶切割(图4)。

本研究发现在水稻细胞质中表达的NifH 蛋白的分子量变小。为解决NifH在植物细胞质中不稳定性问题,对NifH蛋白氨基酸序列进行以系列缺失和氨基酸替代、以及利用烟草烟瞬时表达系统进行研究,结果表明NifH的切割发生在第17位苏氨酸(T)和第18位丝氨基酸(S)之间。13个NifH氨基酸替代突变体(T17A、T17C、T17I、T17M、T17S、T17V、S18A、S18C、S18G、S18L、S18Q、S18T 及 Q19 A),能使 P. polymyxa Δ nifH 突变株恢复固氮酶活性,特别是3个NifH突变体 (T17C、T17V及S18A) 能使Δ nifH 突变株分别恢复到59.4~71.9%固氮酶活性;烟草瞬时表达也证明这13个NifH氨基酸替代突变体在烟草细胞质中不再被蛋白酶切割(图4)。
本研究是国内外首次将13个基因组成的固氮酶合成途径导入水稻中,为实现禾本科植物“自主固氮”提供了重要参考。同时,为预测和解决固氮酶组分及其他原核蛋白在植物细胞质中不稳定性提供有效途径。
我校生物学院陈三凤教授为论文通讯作者,在读博士生尚益民和毕业博士生石皓文(现在工作单位是天津市农业科学院)为论文的共同第一作者。研究得到了“十四五”国家重点研发计划《高效抗逆固氮体系的人工设计与应用示范》项目资助。